History, Evolution, and Social Networks
The encounter of historical and evolutionary perspectives within the intermediate trading zone of social science often has been unsatisfactory. Biological metaphors of social evolution were common among the original founders of the social sciences—in sociology and anthropology especially—but collectivist functionalism1 now is thoroughly discredited. Horrific misuses of biological and evolutionary “scientific theories” by nineteenth- and twentieth-century racist social movements need no recounting. More recently, sociobiology—the analysis of discrete social behaviors and cultural “memes” as if these were genes in evolutionary competition—has gained an enthusiastic following as a sect, but sociobiology is viewed as simplistic and naive by most contemporary social scientists.
Less well known among social scientists, the reverse reception of historicist arguments in evolutionary biology also has been rocky. Stephen Jay Gould is widely known and praised outside of his own subfield, but his arguments are held at arm’s length if not in disdain by his evolutionist peers. Celebrating “historical contingencies” to them seems tantamount to giving up on scientific explanation altogether. Postmodernists in the social sciences and the humanities are willing to take that step, but contemporary evolutionary biologists (including the late Gould himself) have nightmares of creationists and intelligent designers exploiting indeterminacy in evolutionary theory for their own purposes.
With this history and context in mind, Walter Fontana, Member (1999–2000) in the School of Natural Sciences, Walter Powell of Stanford University, and I are initiating a new Social Science Research Council Working Group on History and Evolution in order to develop theoretical and empirical bridges between disciplines not usually in close conversation: evolutionary biology, history, and the social sciences. This initiative builds upon previous collaborations and working groups at the Santa Fe Institute and upon discussions this year at IAS. Why reopen this Pandora’s box of trying to make evolution and history speak to each other? There are motivational and scientific maturity answers to this question.
On the motivational side, the social sciences seem stuck, as Andrew Abbott has long argued,2 in ever reproducing fractal polarities. Methodologically, this is quantitative versus qualitative, but theoretically this is more like methodological individualism versus social constructivism. Instead of contradicting each other, I would argue that both sides need the other to proceed. Methodological individualist theories3 need “actors” to drive their bottom-up deductions about collective behavioral patterns, but “actors” themselves enter from offstage in this approach as axioms, immune to derivation by the theories themselves. Social constructivism strives for the opposite top-down causality—to derive actors and institutions from cultural meaning and interpretation—but “meaning” (or “reflexivity”) axiomatically is presumed in this approach without deriving the material and behavioral patterns to which cognition and language point. In our recent book The Emergence of Organizations and Markets (Princeton University Press, 2012), Powell and I pose this duality as “in the short run, actors make relations, but in the long run, relations make actors.” Instead of juxtaposing the short-run and the long-run sides of this feedback as contradictory, we suggest that their causal interdependence can be seen as a problem of intercalating time scales. The long-run side of this feedback is currently less well understood than is the short-run. Specifically, we suggest that the largest outstanding gap in social science’s collective understanding is our weak processual knowledge about the emergence of “actors”—where do new types of people, organizations, social movements, states, and markets in history come from?
This social-science question about emergent actors is essentially the same as the famous speciation question that Darwin asked about biology long ago in his The Origin of Species. Charles Darwin changed the scientific world with his grand theory of natural selection (which operates on phenotypic variation coming from outside the theory), but he did not answer his original question. Quite rightly in my view, social scientists and even more so historians frequently are repelled by the unilinear progress implications of what they perceive to be Darwinist theory, without appreciating how much of that derives from Herbert Spencer’s “survival of the fittest” bastardization, rather than Darwin’s own “progressive diversification” interpretation.4 Many social scientists also may not realize that contemporary biochemists have come closer toward answering Darwin’s original biological speciation question than did the mid-twentieth-century “Modern Synthesis” of natural selection and genetics into population genetics—that methodologically individualist approach to natural selection based on populations of selfish genes.5
Perhaps the most powerful recent advance in evolutionary biochemistry relevant for this proposal is “evo-devo”—an acronym for evolutionary developmental biology. This is not the study of genes as changing bags of different colored balls, but instead is the study of genes as unfolding regulatory (i.e., feedback) networks of interconnected and sequentially expressed genes and proteins in fetal development. The empirical discovery of a surprisingly low number6 of genes in the human genome has refocused research attention on the combinatorics and sequencing of unfolding gene and protein networks, which actively construct tissues that in turn sequentially interact. Morphological differences among species now seem to be rooted as much in evolving chemical network structures as in evolving genes per se. Evo-devo bridges the gap (or at least tries to) between genotype and phenotype through chemical networks constructing and guiding the interaction of tissues—without denying the competitive selection dynamics of genes-as-inheritance that lie at the heart of all forms of Darwinism.
In a somewhat less momentous way, contemporary evolutionary biology also is being reframed by ecological perspectives, through the concepts of “stigmergy” and “niche construction.” These terms refer to evolutionary feedback between critters (and their social interaction) and the resource environments that those critters themselves partly create. That is, critters make physical environments, which in turn may shape their own evolution if their changes are lasting or repetitive enough. These ecological perspectives gain network twists if “environment” is conceptualized not just as physical resource spaces of consumption but as food webs of multiple species eating each other. Microgenetic (and protein) regulatory networks and macroecological networks must interact because they are all chemical transformations in the end, but coupling network dynamics across multiple time scales is the next research frontier for evolutionary biology.
What does this scientific maturity in biochemistry imply for the social sciences? I propose that history likewise might fruitfully be analyzed as interacting sets of dynamically evolving (and tipping) networks—but of people and practices, not of phenotypes and genotypes.
Fatal missteps of past discussions of biology have followed from slavish social-science efforts to imitate biological reasoning—such as by arguing that biological genes (or pseudogenes like “memes”) should become logical foundations for the social sciences. We social scientists should recognize, however, that biology itself has moved way beyond those primitive notions of unilineal progress and relative species superiority. Once interacting networks replace methodologically individualist genes (or anything else monadic), then coevolution replaces evolution as the outcome to explain. Fixed mountains of optimality dissolve into locally malleable adaptive landscapes that change with movement upon them. Genes and species don’t defeat each other so much as learn (through relative reproduction as well as other mechanisms) how to fit together in mutually consistent (which includes agonistic) ways. Autocatalytic network systems can reproduce themselves through coevolution without being superior or optimal in any global sense.
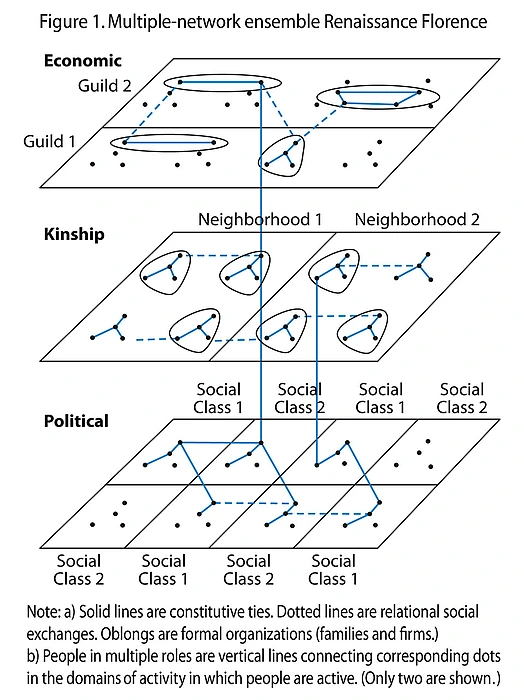
In our recent book, Powell and I have sketched one way in which coevolution and autocatalysis can be conceptualized in terms of multiple social networks—political, economic, kinship, etc.—in dynamic interaction through time. It is not a complete theory as much it is a proof-of-concept that the social speciation question about the emergence of novel actors can be understood in terms of dynamic interaction and cascading feedback among multiple networks. Network analysis already is booming in the social sciences and hardly needs the SSRC to catalyze it. But this subfield has not been placed into dialogue with macroevolutionary and historical questions at the foundations of social science. (Instead network analysis currently is being pushed along more by its synergies with the internet and computer science.) In between the segregated fields of evolutionary biology, history, and network analysis lies a potentially powerful generative space (“structural hole” in the language of Ronald Burt7) where new insights reside, if only mutually respectful conceptual bridges can be built to catalyze them.
What does reframing of evolutionary processes (either biological or social) in terms of networks imply for our understanding of history? Darwin’s idea of biological explanation was not the point predictions of physicists. History to him was a branching bush of diversifying path-dependent trajectories, not a teleological optimum toward which all are heading. The goal of science in this image is not the determinate derivation of the structure of the entire bush; it is the discovery of iterative processual mechanisms that induce both trajectories on the one hand and branching on the other. Discovering mechanisms, not predicting outcomes, is the primary objective of science in its biological variant.8 We now understand better than did Darwin that network features of micromechanisms are responsible for generating nonlinear (and hence hard to predict) macrodynamics. Trajectories (“continuities”) and branching (“change”) are not distinct logics in history according to this biologically inspired view, they are just different phases of common (albeit yet to be discovered) underlying iterative dynamics.
Is this just a more sophisticated nonlinear way of conceptualizing determinism? No. Even within single-network nonlinear systems, bifurcations leave mathematically indeterminate which route to take. Within socially more realistic multiple-network systems, catalytic feedbacks across systems provide one way of conceptualizing “historical contingencies” as consequential interactions between simultaneous parallel processes, which amplify or dampen each other. Surely, this is not the only way of conceptualizing historical contingency, but it is a way mutually respectful of (indeterminate) history, evolutionary biology, and network analysis. The classic problem of agency versus structure thereby gets reframed away from two competing and contradictory logics to a focus on understanding dynamic system responsiveness to various perturbations, including agentic ones.9 Great individuals frequently can be found in exciting case studies, but the science of history resides in understanding how (multiple) networks of others interacted and responded at particular moments in time.10 More generally, social structures become fluid vortexes, not constraining buildings, in a processual understanding.
The activities of the SSRC Working Group on History and Evolution will be three: (a) to convene an inaugural conference of about twenty-five sympathetic social scientists, historians, and evolutionary biologists in order both to explore these themes and to identify committed future participants, (b) to meet regularly as a ten- to twelve-person working group of junior and senior scholars over two to three years in order to develop theory and empirical applications, and (c) to produce an edited SSRC volume that lays out and exemplifies an agenda for the budding field. The SSRC book models I have in mind are Charles Tilly’s Formation of National States in Western Europe (Princeton University Press, 1975) and Theda Skocpol’s Bringing the State Back In (Cambridge University Press, 1985), both landmark examples of SSRC influence on budding social-science agendas. Like these highly successful predecessors, this new SSRC volume will not be a passive summary of existing knowledge and approaches but rather an active intervention, demonstrating how to do interdisciplinary research in an (almost) virgin territory.
Particular sample questions worthy of further exploration, debate, and development by this working group include the following:
1. If genes and memes are insufficient microfoundations for a social-science analogue to contemporary evo-devo, what are the substitutes? Powell and I propose production rules (for production autocatalysis), relational protocols (for biographical autocatalysis), and symbolic addresses (for linguistic autocatalysis). We suggest that interactionally induced assemblages of such practices within people are the disarticulated “under the hood” content of human “agency.” Agency thereby becomes socially diverse, not universally homogeneous, as one corollary. Are these three subsets sufficient to operationalize a practice-based social science, or are more microevolutionary components required? “Consciousness” is a notable omission in the above list, but does that comforting human quality have to appear deus ex machina in our theories, or can it too become an emergent property?
2. If not simplistic “survival of the fittest,” then what is “selection” in a network-based approach? Powell and I propose that “autocatalysis” is the answer, but is that exhaustive of the abstract ways in which practices (of various kinds) differentially reproduce through human interaction? Parasitism and symbiosis, in addition to the usual competition, already are implied within autocatalysis. But mechanisms of multiple-network regulation—where the numerous autocatalyses of overlapping social networks mutually support or undermine each other—remain to be articulated.
3. Both physics and biological natural selection lean heavily on modular architectures11 of units within units (for example, atoms within molecules within gene/protein macromolecules within tissues within organisms within species, etc.) to work. Yet despite the sometime existence of cliques in single networks, multiple-network ensembles of social networks are rarely modular. The cross-cutting overlap of multiple networks is the topological essence of their mutual regulation and coconstitution.12 How modular does the social-science analogue to network-based evo-devo have to be? What do evolution, development, and the genesis of novelty look like in more heterarchical architectures?
4. Political science and sociology colleagues in historical institutionalism recently have become interested in processes of institutional change, in addition to their more traditional focus on comparative equilibria. How does their more macro focus on “institutions” articulate with the more mezzo focus on “networks” articulated here? Obviously, this is a matter of levels of analysis, rather than some deep ontological divide. But how neatly can one segregate time scales, especially in times of historical change when interaction between time scales may be precisely the “spillover” that generates those unintended consequences so frequently observed in empirical case studies about the emergence of novelty?
5. One question that Powell and I get a lot: Does this mean that one has to spend twenty years gathering hard-to-find network data from primary sources before one can have anything to say? The widely diverse (in evidentiary ways as well) empirical chapters in our book are meant to alleviate this career-killing concern. But the fact remains that studying networks, either of social systems or of chemical systems, is data intensive. Biologists have solved this problem through pooling resources on seven or eight “model organisms,” on which most of their biochemical knowledge is based. In the 1960s, SSRC was central in developing organizational models for collaborative research in survey research. Nowadays, efficient computer-search procedures are leading us toward a new world of “big data”—although perhaps at the cost of narrowing social science to the study of Twitter. If the study of “dynamic social networks” is to be more than just the study of the internet, then how can collaboration between social scientists and historians (and any other empirical researchers interested in the deep study of change over time) proceed?
6. If “model organisms” is identified as one useful organizational precedent for coordinating historically dynamic multiple-network research, then what is the set of proof-of-concept empirical cases that we want collectively to study?
7. Powell and I identify “poisedness” as an important next research frontier in the study of emergence. That is, in historical and biological studies of the emergence of “actors,” the macrostudy of the reverberation reaction of networks into which novel actors are inserted is as important as is the microstudy of processual mechanisms of emergence itself. Powell and I describe the emergence of novelty as “innovation”; we call innovation plus network tipping “invention.” If invention is a network system property, then the degree to which previous networks are “on the edge” of tipping becomes important to know. Formal models are suggestive for providing conceptual clues about this question, but empirical historical work, in which “the tape is only played once,” requires disciplined investigation of narrowly constrained counterfactuals to make any progress.
8. “History” is not just the sequential record of events. “History” is also our narration about that record of events. As all historians know, history in the second sense requires authorial selection from the available record of events. History in the first sense, moreover, required selection by whoever coded events into records in the first place. Many historians (including myself) regard the tasteful and argument-driven selection and arrangement of events from records as itself constituting “explanation.”13 Lightly formalizing narrative histories into “narrative networks” of events is one analogical tool for bringing historiographical discussions about alternative narrative representations of the same record into conversation with evolutionary biologists and chemists who also study alternative ways of representing development and evolution.14 Computer scientists who study concurrency in parallel-processing computation could be useful partners in this conversation because they investigate how action rules and timing intercalate. Both evolutionary sequencing in evo-devo and concurrency in parallel-processing computation pose challenges to traditional linear notions of causality in the social sciences similar (or at least analogous) to those posed by historiographical debates about narrative. This topic of the representation of history (and the meaning of causality in such representations) is an important methodological corollary to the substantive question of emergence. Methodological underpinnings need to be addressed in any serious empirical application of theories of nonlinear dynamics in highly interactive systems like multiple networks.
1. Jon Elster, Nuts and Bolts for the Social Sciences (Cambridge University Press, 1989). In contrast, individualist functionalism, in the form of rational choice, is thriving. The definition of “functionalism” used in these sentences is simply “cause explained by consequence.”
2. Andrew Abbott, Chaos of Disciplines (University of Chicago Press, 2001).
3. By this I refer not only to neoclassical rational choice, but also to bounded rationality and cognitive-science approaches like that of Herbert Simon.
4. Symbolized by the branching bush in Darwin’s notebooks.
5. Population genetics does not even conceptualize, much less analyze, phenotype—relying instead on conceptualizing “species” as population-level gene distributions.
6. Lower than any scientist guessed in a lottery before the announcement.
7. Ronald Burt, Structural Holes (Harvard University Press, 1992).
8. This is not to say, however, that short-term prediction does not remainvery important in empirical testing.
9. Like in evolutionary biology: Sure, the important historical contingency of the meteor killed the dinosaurs. But meteors have hit the earth many times without similar consequences. Ecosystem responsiveness to shock at that point in evolutionary time is as central to the explanation of dinosaurs’ extinction as is the meteor itself.
10. Of course in the long run, great men too are made by (iterations of) relations.
11. In his “The Architecture of Complexity,” Proceedings of the American Philosophical Society (Dec 1962), Herbert Simon called this “nearly decomposable systems.” He further made the powerful argument that this architectural feature of observed structure is a deep consequence of the separation (or quantization) of time scales between the multiple dynamic processes that generated it.
12. This idea, of course, is not foreign to biology—nerve systems, blood systems, immune systems, respiratory and digestive systems also overlap and cross-cut in exquisite mutual interaction as well.
13. The word “interpretation” is more frequently used in this context, but equating the two words without getting too hung up on epistemology leads to more fruitful interdisciplinary dialogue than does rigidly insisting on their distinction.
14. Narrative networks are useful for steering between the Scylla of structural determinism and the Charybdis of radical indeterminacy because they make well defined the question of overlap and divergence of multiple representations. See, for example, the intriguing aggregation of fourteen oral histories by Chinese villagers about the “same” Cultural Revolution in their village in Peter Bearman et. al.’s “Blocking the Future,” Social Science History, 1999.